 2015, Vol. 17
2015, Vol. 17



早产儿是指胎龄 <37 周的新生儿。国外研究 发现早产儿骨密度偏低,给新生儿骨胳的生长发 育带来不利的影响,是发生早产儿代谢性骨病最 重要的独立危险因素 [1]。脂联素是由脂肪细胞合成 和分泌的一种蛋白质,调节新生儿的能量摄取, 参与新生儿的生长发育过程 [2]。多种研究发现脂联素可能与其受体结合直接参与骨代谢 [3, 4, 5],而早产 儿脂联素水平和组分发生改变 [6, 7],那么这种改变 与早产儿骨密度水平的降低之间是否存在联系? 目前国内外尚无对血清脂联素与早产儿骨密度关 系的相关研究。本研究采用酶联免疫吸附试验 (ELISA)测定早产儿血清脂联素水平,同时予超 声定量技术评估骨密度,探讨血清脂联素与早产 儿骨密度的关系。
1 资料与方法 1.1 研究对象以 2009 年 3~9 月在我院新生儿专科住院的 1 周内早产儿为研究对象,以同期住院的 1 周内足 月新生儿为对照组,排除中枢神经系统疾病、怀 疑有神经肌肉疾病、接受糖皮质激素治疗及母亲 患有严重妊娠并发症的患儿。根据患儿胎龄分为 3 组:(1)早期早产儿组(31~33+6 周):13 例, 其中男 9 例,女 4 例;(2)晚期早产儿组(34~36+6 周):16 例,其中男 8 例,女 8 例;(3)足月儿 组(37~42 周):43 例,其中男 25 例,女 13 例。 本研究获得新生儿家属的知情同意。
1.2 人体测量学指标测量新生儿出生体重、身长、头围、小腿长 度及二头肌部、三头肌部、髂棘上部和肩胛下角 部的皮脂厚度,采用 Ponderal 指数 [PI=100× 体重 (g)/ 身长(cm)3] 评价新生儿营养状态,采用 Weststrate 公式计算新生儿体脂百分含量。
1.3 骨密度评估使用定量超声仪,根据左胫骨中段超声波在 骨骼中的传播速度(SOS)评估骨密度。
1.4 脂联素的测定新生儿生后 7 d 内于上午 8~10 时采集静脉血 1 mL,不抗凝,室温下放置 2~4 h 充分凝血,然后 以 3 000 r/min 离心 15 min 所得血清置于 -70℃冰 箱保存,使用 ELISA 法测定血清中脂联素浓度。
1.5 统计学分析采用 SPSS 16.0 统计软件进行统计学分析。计 量数据用均数 ± 标准差(x± s)表示,组间比较 采用单因素方差分析,组间两两比较用 SNK-q 检 验。各指标间的相关性采用双变量相关分析和偏 相关分析,并对其相关因素做多元线性逐步回归 分析。 P<0.05 为差异有统计学意义。
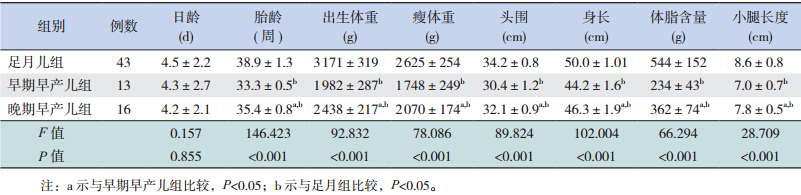
2 结果 2.1 3 组间一般资料的比较早期早产儿组、晚期早产儿组、足月儿组之 间出生体重、瘦体重、头围、身长、体脂含量、 小腿长度的差异均有统计学意义(均 P<0.01), 且随着胎龄增大,出生体重、瘦体重、头围、身长、 体脂含量、小腿长度增加(表 1)。
| 表 1 3 组新生儿一般资料的比较(x± s) |
生 后 1 周 内,早 期 早 产 儿 组 胫 骨 SOS 值 低 于晚期早产儿组和足月儿组,差异有统计学意义 ( P<0.001);晚期早产儿组胫骨 SOS 值低于足月 儿组,差异有统计学意义(P<0.05)(表 2)。
| 表 2 3 组新生儿胫骨 SOS 及血清脂联素的比较(x± s) |
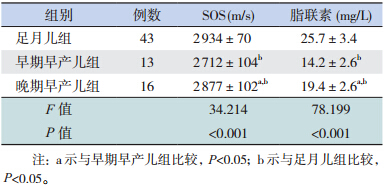
早期早产儿组血清脂联素水平低于晚期早产 儿组和足月儿组,差异有统计学意义(P<0.001); 晚期早产儿组血清脂联素水平低于足月儿组,差 异有统计学意义(P<0.001)(表 2)。
2.4 血清脂联素与生后 1 周内人体测量学指标等的关系生后 1 周内,早产儿血清脂联素水平与出生 体重、胎龄、身长、头围、体脂含量、小腿长度 均呈正相关(分别 r=0.613、0.761、0.504、0.518、 0.624、0.544,均 P<0.05);与日龄、PI 无明显相 关性(P>0.05)。
生后 1 周内,足月儿血清脂联素水平与出生 体重、胎龄、身长、头围、体脂、PI 均呈正相关(分 别 r=0.536、0.720、0.597、0.450、0.565、0.389, 均 P<0.05); 与 小 腿 长 度、 日 龄 无 明 显 相 关 性 (P>0.05)。
2.5 胫骨 SOS 与人体测量学指标等的关系生后 1 周内,早产儿胫骨中段 SOS 值与出生 体重、胎龄、身长、头围、体脂含量、小腿长度、 PI 均呈正相关(分别 r=0.642、0.619、0.445、0.522、 0.524、0.385、0.476,均 P<0.05);与年龄、性别、 钙、磷、钙磷乘积无明显相关性(P>0.05)。
生后 1 周内,足月儿胫骨 SOS 值与出生体重、 胎龄、身长、头围、体脂含量、小腿长度、PI 均 呈正相关(分别 r=0.357、0.628、0.442、0.380、0.477、 0.307、0.308,均 P<0.05),与年龄、性别、钙、磷、 钙磷乘积无明显相关性(P>0.05)。
2.6 血清脂联素与胫骨 SOS 值的关系( r=0.664,P<0.001),调整胎龄、出生体重、体 脂含量、PI、小腿长度后,相关性仍然存在( r=0.451, P=0.031)。
脂 联 素 与 足 月 儿 胫 骨 SOS 值 呈 正 相 关 ( r=0.662,P<0.001),调整胎龄、出生体重、体 脂含量、PI、小腿长度后,相关性仍然存在( r=0.378, P=0.021)。
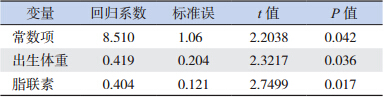
2.7 胫骨 SOS 值的影响因素(1)早产儿胫骨 SOS 值的影响因素:以出生 体重、胎龄、PI、身长、头围、体脂含量、小腿长度、 脂联素为自变量,以早产儿胫骨 SOS 值为因变量, 采用逐步多元线性回归分析上述各因素与胫骨 SOS 值的关系,显示脂联素和出生体重是早产儿胫骨 SOS 值的独立影响因素(表 3)。
| 表 3 早产儿胫骨 SOS 值的独立影响因素 |
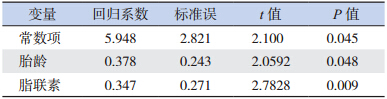
(2)足月儿胫骨 SOS 值的影响因素:以出生 体重、PI、胎龄、身长、头围、小腿长度、体脂含量、 脂联素为自变量,以足月新生儿胫骨 SOS 值为因 变量,采用逐步多元线性回归分析上述各因素与 胫骨 SOS 值的关系,显示脂联素和胎龄是足月儿 胫骨 SOS 值的独立影响因素(表 4)。
| 表 4 足月儿胫骨 SOS 值的独立影响因素 |
新生儿时期的骨状况对人体有重要的影响, 新生儿时期的骨骼发育是成人骨骼的基础,胎儿 和婴儿早期骨营养状况可以影响人体骨的整个生 长发育过程 [8]。有报道显示早产儿的骨矿化持续 落后 [9]。因此客观地评价早产儿骨的营养状况非 常重要。近来很多研究都表明,骨定量超声技术 可用来评估早产儿骨骼状态 [10]。骨定量超声不但 可测量骨矿物质密度、骨皮质厚度、弹性,而且 还可以显示骨微结构。因此本研究选用骨定量超 声技术测定早产儿和足月儿胫骨 SOS 值来评估骨 密度,发现不同胎龄组间胫骨 SOS 值有差异,其 中早期早产儿组胫骨 SOS 值低于晚期早产儿组和 足月儿组,晚期早产儿组胫骨 SOS 值低于足月儿 组,差异均具有统计学意义。与 Ashmeade 等 [11]、Tomlinson 等 [12]、刘晓莉等 [13] 的研究结果一致。早 产儿,尤其是极低出生体重儿出生时骨密度明显 低于足月儿,到纠正胎龄足月时仍多不能赶上健 康足月儿水平,而且这种差别可持续到生后 6 个 月 [14],如合并早产儿代谢性骨病者,骨的低密度 可持续至出生 1~2 岁 [15]。新生儿骨密度与胎龄、 体重密切相关,因为胎儿后期是骨矿物质储备的 主要时期,由于早产儿过早分娩,从母体获得的 骨矿物质不足,出生时体内储备量较少,胎龄越小, 体重越低,宫内矿物质沉积越少,因此骨密度低 于足月儿。
脂联素是近年来发现的一种由脂肪组织分泌 的蛋白质类激素,具有促进脂质氧化、调节能量 代谢平衡、参与骨的代谢活动等多种生物学作用, 在胎儿生长发育中起着重要的作用。本研究结果 显示,新生儿血清脂联素水平在不同胎龄分组间 差异有统计学意义,早期早产儿组脂联素水平低 于晚期早产儿组和足月儿组,晚期早产儿组脂联 素水平低于足月儿组,且相关分析显示血清脂联 素水平与出生体重、胎龄、头围、身长及体脂含 量等均呈正相关,提示新生儿血清脂联素水平与 宫内的生长发育密切相关,随着胎龄和体重的增 加,血清脂联素浓度也增高。国内董红红等 [16] 测 定不同胎龄新生儿出生 1 d 时的血清脂联素水平, 结果显示早产儿组脂联素水平明显低于足月儿组, 且与胎龄呈正相关,随着胎龄的增加呈上升趋势, 与国外学者的研究结果一致 [17],提示脂联素参与 新生儿的生长发育过程。脂联素在促进骨代谢方 面亦具有积极作用,目前认为脂联素通过内分泌、 自分泌和旁分泌 3 个不同的途径影响成骨细胞及 破骨细胞的活性,积极参与骨代谢 [18]。Luo 等 [19] 研究脂联素对人成骨细胞增殖和分化的分子机制, 研究发现成骨细胞是脂联素的直接靶细胞,脂联 素诱导人成骨细胞增殖和分化。同时,脂联素通 过 MAPK 途径诱导人类成骨细胞破骨细胞分化因 子(RANKL)表达,抑制护骨素(OPG)的表达, 间接诱导破骨细胞分化。体外研究表明,脂联素 通过抑制破骨过程和激活成骨过程,从而增加骨 矿含量,发挥提高骨量的活性 [20]。不同部位的脂 联素影响骨形成的途径不同。局部产生的脂联素 可能通过内分泌或旁分泌途径促进骨形成,而循 环中脂联素既可直接抑制骨形成,又可通过促进胰岛素信号转导间接促进骨形成。
国内外尚无关于脂联素与新生儿骨密度关系 的研究。脂联素与青春期和成年期骨密度关系的 研究较少,目前尚无一致的结论。Russell 等 [21] 研 究发现脂联素与 12~18 岁青春期女性的全身骨密 度呈负相关,内脏脂肪是全身骨密度的独立负性 决定因素。台湾的一个研究发现血清脂联素水平 与青春期女性全身骨密度呈负相关,调整年龄、 BMI、全身体脂含量后相关性不存在 [22]。脂联素与 绝经后的 41~81 岁中国女性的骨密度呈负相关, 脂联素可能通过促进绝经后妇女的骨吸收对其骨 密度施加负面影响 [23]。李莉军等 [24] 的研究发现脂 联素与各部位骨密度显著相关,校正年龄、体脂 后,脂联素与各部位骨密度相关性存在;进一步 逐步多元线性回归分析显示脂联素是男性各部位 骨密度的独立影响因素。本研究发现,血清脂联 素水平与早产儿和足月儿骨密度均呈正相关,脂 联素和出生体重是早产儿骨密度的独立影响因素, 而脂联素和胎龄是足月儿骨密度的独立影响因素。 本研究结果与成年期及青春期相关研究不同,可 能与新生儿期脂联素受体的分布和密度与青春期 和成年期不同有关,新生儿期成骨细胞表面脂联 素受体的密度远远大于破骨细胞表面的受体密度, 因此脂联素对骨骼发育的作用就主要表现为成骨 作用;而青春期和成年期主要是破骨细胞表面脂 联素受体的密度较高,脂联素对骨骼的影响就表 现为破骨作用或二者作用抵消。另一方面,早产 儿脂联素的构成发生了改变 [6, 7],不同分子量的脂 联素作用不同,可能影响脂联素的成骨作用;新 生儿期脂联素的来源与青春期和成年期不同,新 生儿期脂联素除来源于白色脂肪组织外,还来源 于棕色脂肪组织,不同来源的脂联素可能发挥的 生物学效应也不同。总之,脂联素对早产儿骨骼 发育影响的具体机制仍需进一步的研究。
| [1] | Hovi P, Andersson S, Järvenpää AL, et al. Decreased bone mineral density in adults born with very low birth weight: a cohort study[J]. PLoS Med, 2009, 6(8): e1000135. |
| [2] | 董红红, 李彦敏, 田朝霞, 等. 早产儿血清脂联素水平测定 的临床意义[J]. 实用儿科临床杂志, 2009, 24(14): 1807. |
| [3] | 袁凌青, 罗湘杭, 谢辉, 等. 脂联素对人成骨细胞增殖作用 的研究[J]. 中国骨质疏松杂志, 2006, 12(2): 135-137. |
| [4] | Luo XH, Guo LJ, Yuan LQ, et al. Adiponectin stimulates human osteoblasts proliferation and differentiation via the MAPK signaling pathway[J]. Exp Cell Res, 2005, 309(1): 99-109. |
| [5] | Luo XH, Guo LJ, Xie H, et al. Adiponectin stimulates RANKL and inhibits OPG expression in human osteoblasts via the MAPK signaling pathway[J]. J Bone Miner Res, 2006, 21(10): 1648-1656. |
| [6] | Yoshida T, Nagasaki H, Asato Y, et al. The ratio of highmolecular weight adiponectin and total adiponectin differs in preterm and term infants[J]. Pediatr Res, 2009, 65(5): 580-583. |
| [7] | Siahanidou T, Margeli A, Garatzioti M, et al. Disparity in circulating adiponectin multimers between term and preterm infants[J]. J Perinat Med, 2009, 37(6): 683-688. |
| [8] | Korakaki E, Damilakis J, Gourgiotis D, et al. Quantitative ultrasound measurements in premature infants at 1 year of age: the effects of antenatal administered corticosteroids[J]. Calcif Tissue Int, 2011, 88(3): 215-222. |
| [9] | Zamora SA, Belli DC, Rizzoli R, et a1. Lower femoral neck bone mineml density in prepubertal former preterm girls[J]. Bone, 2001, 29(5): 424-427. |
| [10] | 赵琳, 李琳霞, 李艳红. 骨定量超声技术评价早产儿骨量 和早产儿骨发育的影响因素[J]. 中国新生儿科杂志,2012, 27(3): 183-185. |
| [11] | Ashmeade T, Pereda L, Chen M, et al. Longitudinal measurements of bone status in preterm infants[J]. J Pediatr Endocrinol Metab, 2007, 20(3): 415-424. |
| [12] | Tomlinson C, McDevitt H, Ahmed SF, et al. Longitudinal changes in bone health as assessed by the speed of sound in very low birth weight preterm infants[J]. J Pediatr, 2006, 148(4): 450-455. |
| [13] | 刘晓莉, 白增华. 早产儿和足月儿骨营养状况的分析研究[J]. 中国优生与遗传杂志, 2010, 18(5): 132-135. |
| [14] | 杨文旭, 杨慧明. 早产婴儿骨密度及其影响因素分析[J]. 中 国当代儿科杂志, 2011, 13(11): 859-862. |
| [15] | 张荣娜, 修文龙. 早产儿代谢性骨病[J]. 实用儿科临床杂志, 2011, 26(8): 624-627. |
| [16] | 董红红, 田朝霞, 娜琪, 等.早产儿血清脂联素水平及其与 血脂关系的研究[J]. 中国新生儿科杂志, 2012, 27(1): 40-41. |
| [17] | Lecke SB, Morsch DM, Spritzer PM. Leptin and adiponectin in the female life course[J]. Braz J Med Biol Res, 2011, 44(5): 381-387. |
| [18] | 赵勤, 罗湘杭. 骨代谢过程中脂联素的相关作用[J]. 中国组 织工程研究与临床康复, 2007, 11(6): 1127-1129. |
| [19] | Luo XH, Guo LJ, Yuan LQ, et a1. Adiponectin stimulates human osteoblasts proliferation and diferentiation via the MAPK signaling pathway[J]. Exp Cel Res, 2005, 309(1): 99 -109. |
| [20] | 郝东升, 黎明. 脂联素与骨代谢的研究进展[J]. 中华骨质疏 松和骨矿盐疾病杂志, 2009, 2(4): 260-267. |
| [21] | Russell M, Mendes N, Miller KK, et al. Visceral fat is a negative predictor of bone density measures in obese adolescent girls[J]. J Clin Endocrinol Metab, 2010, 95(3): 1247-1255. |
| [22] | Huang KC, Cheng WC, Yen RF, et al. Lack of independent relationship between plasma adiponectin, leptin levels and bone density in nondiabetic female adolescents[J]. Clin Endocrinol (Oxf), 2004, 61(2): 204-208. |
| [23] | Zhang H, Xie H, Zhao Q, et al. Relationships between serum adiponectin, apelin, leptin, resistin, visfatin levels and bone mineral density, and bone biochemical markers in postmenopausal Chinese women[J]. J Endocrinol Invest, 2010, 33(10): 707-711. |
| [24] | 李莉军, 郭丽娟, 孙文莉. 男性血清抵抗素、脂联素水平与 骨密度关系[J]. 中国骨质疏松杂志, 2008, 14(1): 47-49. |