 2015, Vol. 17
2015, Vol. 17



2. 四川大学华西公共卫生学院 营养与食品卫生学及毒理学系, 四川 成都 610041;
3. 四川大学华西第二医院产科, 四川 成都 610041
双歧杆菌是人体肠道里最早出现的肠道微生 物之一[1],是婴儿肠道微生物的优势菌,也是人体 肠道内最主要的有益菌之一,对维持人体健康有 着极其重要的作用。双歧杆菌不仅具有营养功能,如产生维生素,同时还具有抗感染、抗癌作用; 双歧杆菌通过促进肠道黏膜 SIgA 系统的成熟等机 制促进婴儿免疫系统的发育和成熟[2, 3, 4]。因此,双 歧杆菌的正常发育,即在合适的时间定植合适的 双歧杆菌,对婴儿的健康成长有着重要的意义。
双歧杆菌的定植受很多因素的影响,如生产 方式、喂养方式、孕周、抗生素的使用、环境地 域等[5]。国内对婴儿早期肠道双歧杆菌的定植情况 及动态变化的相关研究很少,且双歧杆菌检测种 类少、检测时间点选择代表性不够,在婴儿双歧 杆菌的定植方式及特点上还存在许多不明的地方。 为了解婴儿肠道双歧杆菌群构建的规律,本研究 首次采用分离培养法结合分子生物学方法,对婴 儿早期肠道中双歧杆菌种群构造进行了全面系统 的分析,拟为进一步深入探讨双歧杆菌对婴儿健 康的影响提供有益的基础数据。 1 资料与方法 1.1 研究对象
选取2013 年3~4 月间于四川大学华西第二医 院出生的正常足月新生儿为观察对象。纳入标准: 居住在成都市五城区;胎龄37~42 周;出生体重 2 500~4 000 g;无先天性异常或出生缺陷。排除标 准:母亲产前1 个月内使用过抗生素;父母有结 核病、乙肝、艾滋病等传染性疾病;母亲有胃肠 道疾病;母亲患有重大疾病;新生儿因病理性黄疸, 新生儿肺炎等疾病服用抗生素或微生态制剂。本 研究遵循Helsink 原则,由纳入研究的新生儿父亲 或母亲签署知情同意书。共募集到28 名孕妇生产 的30 例新生儿(包括两对双胞胎),其中16 例 完成3 个月追踪调查纳入本研究:包括男7 例, 女9 例;剖腹产13 例,顺产3 例;母乳喂养7 例, 混合喂养9 例。另外14 例新生儿(包括两对双胞胎) 因新生儿疾病或其他原因退出调查。 1.2 材料和试剂
粪便稀释液的配制: 将4.5 g KH2PO4、 6.0 g Na2HPO4、0.5 g L- 半胱氨酸盐酸盐、0.5 g Tween-80 和1.0 g 琼脂混合于1 000 mL 蒸馏水中, 121 ℃ 20 min 高压灭菌;BL 培养基(BL agar) 和TOS 培养基(TOS Propionate agar) 为日本荣 研化学株式会社产品;MRS 培养基为北京陆桥公 司产品;厌氧产气袋为日本三菱化学株式会社产 品;双歧杆菌标准菌株购于日本理化学研究所微 生物保存施设(Japan Collection of Microorganism, JCM): 青春双歧杆菌(Bifidobacterium adolescentis,B. adolescentis)ATCC15703、长 双歧杆菌(Bifidobacterium longum,B. longum) ATCC15707、短双歧杆菌(Bifidobacterium breve, B. breve)ATCC15700、婴儿双歧杆菌 (Bifidobacterium infantis,B.infantis) ATCC15697、两歧双歧杆菌(Bifidobacterium bifidum,B. bifidum)ATCC29521、链状双歧杆菌 (Bifidobacterium catenulatum,B. catenulatum) ATCC27539、角双歧杆菌(Bifidobacterium angulatum,B. angulatum)ATCC27535、齿双 歧杆菌(Bifidobacterium dentium,B. dentium) ATCC27534;2×Taq MasterMix( 含染料) 为北 京康为世纪科技有限公司产品;引物合成由大连 TaKaRa 公司完成;PCR 仪、电泳仪及紫外凝胶成 像系统均购自美国BIO-RAD 公司。 1.3 一般资料及样本采集
对纳入的对象进行问卷调查(包括分娩方式、 喂养方式、婴儿性别、婴儿健康状况及用药情况 等)。分别收集婴儿出生后第1 次、第2、4、7、 10、14、28、90 天共8 次粪便于单独灭菌的螺旋 式粪便盒。采样后立即送检,如不能立即检测, 则厌氧4℃保存,当日检测。 1.4 双歧杆菌的分离
采用文献[6] 的方法,对采集的粪便中双歧杆 菌进行分离培养。称取粪便0.5 g,加入4.5 mL 的 粪便稀释液,振荡混匀,并作10 倍系列稀释。 取适宜浓度的混合液各100 μL,均用L 型玻璃棒 均匀涂布于BL 平板(含5% 马血),37℃厌氧 培养48~72 h。根据双歧杆菌在非选择性培养基 BL 平板上生长的独有特征(中央棕色且边缘部分 呈棕红色,圆形,隆起,边缘整齐,表面光滑湿 润,直径3~5 mm),从最高稀释倍数的BL 平板 上,分别挑取具有不同特征的可疑菌落接种于双 歧杆菌选择性培养基TOS 平板,置于37℃厌氧 培养48 h。挑取TOS 平板上的细菌进行革兰染色 并在显微镜下观察细菌形态,镜检为革兰阳性杆 菌,着色不均,弯曲,一端或两端膨大、分叉, 无芽孢且接种于MRS 平板,37℃需氧培养48 h 后细菌严格厌氧的菌株,初步判定为双歧杆菌,用 11%~12% 灭菌的脱脂奶对菌株进行保存。 1.5 双歧杆菌的PCR 鉴定
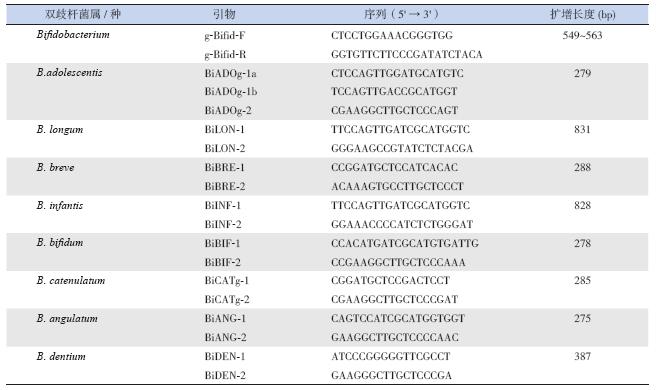
用煮沸法提取细菌DNA:从平板培养基上 挑取适量细菌于100 μL TE Buffer(10 mM Tris- HCl,1 mM EDTA,pH=8.0),95 ℃ 加热5 min, 10 000 r/min 离心3 min,吸取上清液,即为细菌 DNA。根据文献[7],合成双歧杆菌属及8 种不同的 双歧杆菌种引物(表 1)。PCR 扩增条件:94℃预 变性2 min;94℃变性30 s,55℃退火30 s,72℃ 延伸30 s,35 个循环;72℃终延伸2 min。PCR 扩 增产物的检测采用琼脂糖凝胶电泳法。
| 表 1 双歧杆菌属/ 种PCR 特异性引物 |
采用SPSS 16.0 统计软件对数据进行统计学分 析,计数资料用百分率表示,组间比较采用卡方 检验或Fisher 确切概率法,P<0.05 为差异有统计 学意义。 2 结果 2.1 婴儿双歧杆菌的定植时间
16 例新生儿的第1 次、第2 天粪便中均未检 出双歧杆菌;第4、7 天分别采集到13 例新生儿 粪便,双歧杆菌定植率均为8%(1/13);随即双 歧杆菌迅速定植,第28 天时共采集到13 例新生 儿粪便,双歧杆菌定植率增至54%(7/13),与第 4 天定植率相比,差异有统计学意义(P=0.03); 3 月龄时共采集到15 例婴儿粪便,双歧杆菌定 植率达到60%(9/15),明显高于第4 天定植率 (P=0.006)。检出双歧杆菌的粪便里,双歧杆菌 的数量为1.0×109~2.0×1011 CFU/g 粪便(双歧杆 菌检出限≥ 105 CFU/g 粪便)。 2.2 婴儿双歧杆菌菌种的变化
16 例婴儿的双歧杆菌菌种构成及变化分析结 果如图 1 所示。婴儿肠道内双歧杆菌菌种由最开 始单一的B. bifidum 逐步演变为多个菌种并存, 随着时间推移逐渐检出B. catenulatum、B. breve、 B. infantis、B. longum、B. dentium 及B. angulatum, 未检出B. adolescentis。成人型双歧杆菌 B. catenulatum 定植时间早于婴儿型双歧杆菌 B. infantis。第14 天时首次出现B. infantis,数量为1.0×108 CFU/g 粪便,而3 个月时B. infantis 则 增长到1.0×109~1.1×1011 CFU/g 粪便。
 |
图 1 婴儿肠道双歧杆菌定植种类随时间的变化 |
婴儿个体之间双歧杆菌定植的种类差异较大。 至3 月龄时,除了3 例婴儿未检出双歧杆菌外, 10 例婴儿肠道中定植的双歧杆菌种类总数不超过 2 种,仅3 例婴儿双歧杆菌种类总数超过2 种,最 多为4 种。因婴儿双歧杆菌定植不稳定,个别婴 儿肠道双歧杆菌在不同的时间点出现种类交替, 16 例婴儿中,6 例出现过2 种及以上双歧杆菌定植, 最早1 例发生在生后第14 天,有4 例在生后第 28 天被检出,1 例在3 月龄时被检出。 2.3 双歧杆菌菌种组成
从婴儿肠道分离鉴定的双歧杆菌共152 株, 除B. adolescentis 未检出外,双歧杆菌菌种为 B. breve 34 株(22.4%)、B. longum 31 株(20.4%)、 B. bifidum 21 株(13.8%)、B. catenulatum 19 株 (12.5%)、B. infantis 15 株(9.9%)、B. dentium 15 株(9.9%)、B. angulatum 2 株(1.3%),其他 15 株(9.9%)。 2.4 喂养方式和生产方式对婴儿双歧杆菌定植的影响
生后第10 天时9 例混合喂养新生儿中只有 1 例(11%)检出双歧杆菌,而同期7 例母乳喂养 新生儿中有3 例(43%)检出双歧杆菌定植;到3 个月时,混合喂养儿中已有5 例(56%)婴儿定植 双歧杆菌,母乳喂养儿有4 例(57%)检出双歧杆 菌。3 例顺产婴儿中,有1 例(33%)至3 月龄仍 未检出双歧杆菌,2 例(67%)仅检出一种双歧杆菌, 分别为B. bifidum 和B. infantis;13 例剖腹产婴儿 中,在生后10 d 时已有4 例(31%)检出双歧杆菌, 28 d 时有7 例(54%)检出双歧杆菌定植,并由单 一的B. bifidum 演变为多个菌种并存,仅1 例(8%) 剖腹产婴儿至3 月龄仍未检出双歧杆菌。 3 讨论
双歧杆菌是婴儿(尤其是母乳喂养儿)肠道 菌群中的优势菌群,可合成维生素,减缓过敏性 皮炎的症状,减少腹泻、病毒性感染并减轻乳糖 不耐受,尤其在婴儿免疫功能发育方面起着不容 忽视的作用[2]。目前为止的研究表明,双歧杆菌作 为婴儿肠道首批定植的细菌之一,紧随在葡萄球 菌、链球菌和肠杆菌等需氧菌或兼性厌氧菌定植 之后,是婴儿肠道内定植最早的专性厌氧菌之一。 婴儿出生后数小时内,双歧杆菌即开始在肠道内 定植,并迅速增殖(尤其是母乳喂养儿);出生 一周后双歧杆菌大量增殖;出生一个月左右婴儿 肠道菌群结构发生显著变化,3 月龄时双歧杆菌成 为婴儿肠道优势菌群[8]。
而国内对婴儿早期肠道双歧杆菌定植和发 展的研究很少,检测的种类也局限于B. breve、 B. longum 等亚种[9],且双歧杆菌检测时间点不够 系统[9, 10]。本研究通过对受试婴儿粪便中分离培养 到的双歧杆菌进行菌种水平的鉴定,首次对婴儿 早期肠道中双歧杆菌种群构造进行了全面系统的 分析。
本研究中,受试婴儿出生4 d 粪便开始有双 歧杆菌定植,且数量可达到6.0×109 CFU/g 粪便, 定植率为8%。一周后定植率由8% 增至生后28 d 时的54%,至生后3 个月时达到60%,且双歧杆 菌数量维持在较高水平(1.0×109~2.0×1011 CFU/g 粪便)。但是婴儿个体之间双歧杆菌开始定植的 时间差异较大,且双歧杆菌定植不稳定,而且肠 道内已定植双歧杆菌的婴儿个体之间双歧杆菌数 量也存在较大差异。本研究中有3 名婴儿(包括 1 名顺产婴儿)至3 月龄时均未检出双歧杆菌,这 同Azad 等[11] 的研究结果相似,个别婴儿4 个月时 粪便中仍未检出双歧杆菌,包括纯母乳喂养的顺 产婴儿。
大量研究已证明,双歧杆菌是肠道内微生物 里对人体健康有益的代表性菌群之一,维持及提 高双歧杆菌在人体肠道内的数量已被广泛认为是 促进人体健康的重要手段之一。近年来,人体肠 道双歧杆菌除了其数量以外,双歧杆菌的菌种构 成及其多样性对人体健康的影响同样受到关注。 研究表明,双歧杆菌的生理机能具有显著的菌种特异性,双歧杆菌的菌种构成及多样性上的差异 被发现是过敏症等疾患的病理学特征之一[12]。本 研究中,首先出现在受试婴儿粪便中的肠道双歧 杆菌是B. bifidum,随着时间变化双歧杆菌的菌种 组成多样性明显增加。出生10 d 后,受试婴儿肠 道双歧杆菌由单一的B. bifidum 优势菌种发展为 B. catenulatum、B. breve、B. longum、B. dentium、 B. angulatum 以及B. infantis。本研究结果再度表 明B. bifidum 是研究受试婴儿早期最常分离到的双 歧杆菌优势菌种之一。
喂养方式是影响婴儿肠道菌群早期构建的重 要因素[5]。已有研究表明和母乳喂养儿相比,人工 喂养儿双歧杆菌的定植时间延迟[13]。本研究中婴 儿均为母乳喂养或者混合喂养,研究结果提示母 乳喂养婴儿双歧杆菌的定植可能比混合喂养儿早, 但至3 月龄时基本相平。因受试婴儿较少,特别 是本研究此时还没有纯人工喂养的婴儿参加,因 此还不能就喂养方式对婴儿双歧杆菌定植的影响 进行统计分析,将在后续研究中不断完善。
分娩方式是影响婴儿肠道菌群定植的重要因 素之一[5]。和顺产婴儿相比,剖腹产婴儿肠道双歧 杆菌的数量较低,且双歧杆菌定植的种类更少[14]; 剖腹产婴儿双歧杆菌定植延迟,双歧杆菌在1 月 时定植率才达到顺产水平[15]。本研究中,因顺产 婴儿较少,还不能对顺产和剖腹产婴儿双歧杆菌 数量及种类进行统计分析,但考虑到受试婴儿多 数是剖腹产婴儿,因此推测剖腹产可能是造成受 试婴儿双歧杆菌定植延迟的因子之一,但仍需进 一步研究以证实。
Avershina 等[16] 研究表明,婴儿出生10 d 内 的双歧杆菌是由B. longum 和B. breve 为优势菌种 的多菌种所组成。相比之下,本研究的受试婴儿肠 道双歧杆菌的早期多样性显著低于目前为止的研 究报道。考虑到本研究的受试对象多数是剖腹产 婴儿,推测剖腹产可能是造成肠道双歧杆菌早期 多样性减少的原因之一。但是,本研究的受试婴 儿数量整体偏少,今后还要继续观察。本研究结 果和Tsuji 等[17] 的研究基本一致,即在3 个月时本 研究受试婴儿肠道双歧杆菌为肠道优势菌群,包括 B. longum、B. breve、B. infantis 和B. catenulatum。 B. breve、B. infantis 及B. longum 是婴儿肠道最常 见的双歧杆菌,而B. catenulatum、B. adolescentis 及B. longum 是成人粪便中最常分离到的双歧杆 菌[18]。本研究受试婴儿肠道中B. infantis 出现较 晚,在3 个月时才在婴儿粪便中检测到高水平的 B. infantis,显著迟于其他报道[19]。研究报告指出, 患过敏性疾病的婴儿肠道里会出现B. adolescentis 为代表的成人型双歧杆菌[11]。本研究中,虽然未 检出B. adolescentis,但是婴儿肠道第10 天就开始 检出成人型双歧杆菌B. catenulatum,且之后检出 率也较高,这是否和剖腹产有关联,还需要进一 步研究。
本研究通过对婴儿肠道双歧杆菌进行连续全 面的追踪分析,发现受试婴儿早期肠道双歧杆菌 多样性偏低,婴儿型双歧杆菌出现偏迟,相反成 人型双歧杆菌发现较早。上述结果对发现婴儿肠 道双歧杆菌定植的特点和规律奠定了基础,有利 于进一步深入探讨双歧杆菌与婴儿健康的关系。
| [1] | Rautava S, Luoto R, Salminen S, et al. Microbial contact during pregnancy, intestinal colonization and human disease[J]. Nat Rev Gastroenterol Hepatol, 2012, 9(10): 565-576. |
| [2] | Sjogren YM, Tomicic S, Lundberg A, et al. Influence of early gut microbiota on the maturation of childhood mucosal and systemic immune responses[J]. Clin Exp Allergy, 2009, 39(12): 1842-1851. |
| [3] | Holscher HD, Czerkies LA, Cekola P, et al. Bifidobacterium lactis Bb12 enhances intestinal antibody response in formula-fed infants: a randomized, double-blind, controlled trial[J]. JPEN J Parenter Enteral Nutr, 2012, 36(1 Suppl): 106S-117S. |
| [4] | 朱海娟, 蔡燕. 口服双歧杆菌对极低出生体重儿免疫功能的 影响[J]. 中国当代儿科杂志, 2011, 13(12): 944-946. |
| [5] | Penders J, Thijs C, Vink C, et al. Factors influencing the composition of the intestinal microbiota in early infancy[J]. Pediatrics, 2006, 118(2): 511-521. |
| [6] | Mitsuoka T. A color atlas of anaerobic bacteria[M]. Tokyo: Sobunsya, 1980: 341. |
| [7] | Matsuki T, Watanabe K, Fujimoto J, et al. Quantitative PCR with 16S rRNA-gene-targeted species-specific primers for analysis of human intestinal bifidobacteria[J]. Appl Environ Microbiol, 2004, 70(1): 167-173. |
| [8] | Roberts AK, Chierici R, Sawatzki G, et al. Supplementation of an adapted formula with bovine lactoferrin: 1. Effect on the infant faecal flora[J]. Acta Paediatrica, 1992, 81(2): 119-124. |
| [9] | 张红波, 陆薇, 黄娟, 等. 不同分娩方式对母乳喂养婴儿肠 道菌群的影响[J]. 临床儿科杂志, 2010, 28(3): 251-254. |
| [10] | 黄永坤, 曹志琅, 杨武, 等. 出生早期新生儿肠道双歧杆菌 动态检测[J]. 临床儿科杂志, 2010, 28(10): 938-941. |
| [11] | Azad MB, Konya T, Maughan H, et al. Gut microbiota of healthy Canadian infants: profiles by mode of delivery and infant diet at 4 months[J]. CMAJ, 2013, 185(5): 385-394. |
| [12] | He F, Ouwehand AC, Isolauri E, et al. Comparison of mucosal adhesion and species identification of bifidobacteria isolated from healthy and allergic infants[J]. FEMS Immunol Med Microbiol, 2001, 30(1): 43-47. |
| [13] | Favier CF, de Vos WM, Akkermans ADL. Development of bacterial and bifidobacterial communities in feces of newborn babies[J]. Anaerobe, 2003, 9(5): 219-229. |
| [14] | Biasucci G, Benenati B, Morelli L, et al. Cesarean delivery may affect the early biodiversity of intestinal bacteria[J]. J Nutr, 2008, 138(9): 1796S-1800S. |
| [15] | Gronlund MM, Lehtonen OP, Eerola E, et al. Fecal microflora in healthy infants born by different methods of delivery: permanent changes in intestinal flora after cesarean delivery[J]. J Pediatr Gastroenterol Nutr, 1999, 28(1): 19-25. |
| [16] | Avershina E, Storro O, Oien T, et al. Bifidobacterial succession and correlation networks in a large unselected cohort of mothers and their children[J]. Appl Environ Microbiol, 2013, 79(2): 497-507. |
| [17] | Tsuji H, Oozeer R, Matsuda K, et al. Molecular monitoring of the development of intestinal microbiota in Japanese infants[J]. Benef Microbes, 2012, 3(2): 113-125. |
| [18] | Matsuki T, Watanabe K, Tanaka R, et al. Distribution of bifidobacterial species in human intestinal microflora examined with 16S rRNA-gene-targeted species-specific primers[J]. Appl Environ Microbiol, 1999, 65(10): 4506. |
| [19] | Mullie C, Romond MB, Izard D. Establishment and follow-up of bifidobacterial species in the gut of healthy bottle-fed infants of 1-4 months age[J]. Folia Microbiol, 2006, 51(5): 473-477. |