 2016, Vol. 18
2016, Vol. 18


儿童哮喘是一种常见的易反复发作的呼吸道疾病,在全世界范围内分布,深刻影响儿童的健康和生活状态,亦常给患儿家庭带来难以承担之经济及精神负担。目前,哮喘的发病机制尚不清楚,普遍认为是由多种免疫细胞(淋巴细胞、中性粒细胞、嗜酸性细胞等)和多种免疫因子(IL-6、IL-8等)介导的免疫性疾病。IL-17A是IL-17家族中一员,主要由Th17细胞产生,在NKT细胞、γδT细胞、中性粒细胞、嗜酸性粒细胞中也都有表达。已有研究表明,在哮喘患者的血清中IL-17A的表达是明显升高的[1]。而IL-17A基因启动子区的-197G/A和-692C/T位点的多态性与多种免疫性疾病的易感性相关[2]。通过对IL-17A基因-197位、-692位的基因型及等位基因分布频率的测定,本研究拟阐明两位点基因多态性与儿童哮喘易感性的关系。
1 资料与方法 1.1 研究对象随机选取2013年8月至2015年8月于西南医科大学附属医院门诊随访或住院的哮喘患儿65例为研究对象(哮喘组),其中男39例,女26例,平均年龄4.3±2.2岁。所有病例均符合诸福棠实用儿科学关于儿童支气管哮喘的诊断标准[3]。另选取同期行健康体检儿童70例为健康对照组,其中男38例,女32例,平均年龄3.8±2.3岁。入选对照组儿童需近期无发热、感染、过敏、使用激素病史,排除肿瘤、自身免疫性疾病。本研究项目获得西南医科大学附属医院伦理委员会批准并取得患儿及其家属知情同意。
1.2 DNA提取及引物合成采集两组儿童外周静脉血1~2 mL于抗凝管中,取200 μL抗凝血应用DNA提取试剂盒(北京天根生物科技有限公司)提取人基因组DNA;而同时需通过人基因库已知的基因序列及IL-17A两位点的单核苷酸多态性(single nucleotide polymorphism, SNP)设计序列特异性引物并由上海生工合成备用。见表 1。
| 表 1 IL-17A序列特异性引物 |

1.3 序列特异性引物聚合酶链反应
序列特异性引物聚合酶链反应(sequence specific primers-polymerase chain reaction, SSP-PCR)反应体系为25 μL:模板DNA 2 μL、2×Power Taq Master Mix 12.5 μL、特异性正向引物F1及F2各0.5 μL、内参正向引物C 0.5 μL、通用反向引物0.5 μL、ddH2O 8.5 μL。反应条件:94℃预变性4 min;94℃变性30 s,退火(温度见表 1)40 s,72℃延伸40 s,共35个循环;72℃再延伸5 min。
1.4 SSP-PCR产物电泳分析在每个1.5%琼脂糖凝胶样孔中分别加入PCR产物5 μL,再将凝胶放入室温状态下电压为110 V的电泳仪(0.5×TAE缓冲液)中电泳35 min,最后将电泳后的凝胶置于自动凝胶成像仪(基因公司,英国)里查看电泳图谱并记录结果。
1.5 PCR产物测序通过以上实验流程,初步确定每例研究对象SNP位点的基因型。随机选取两位点各3份DNA样本(均为杂合子)用内参引物(C,R)进行PCR扩增,产物送至上海生工进行DNA测序。测序结果与应用SSP-PCR法检测的SNPs位点的基因型结果比对,以此验证SSP-PCR法检测SNPs的可靠性和准确性。
1.6 统计学分析采用SPSS 18.0统计软件对数据进行统计学分析。基因型及等位基因的分布频率采用百分率(%)表示,两组间比较采用卡方检验;相对危险度采用优势比(OR)评估,同时计算95%可信区间(CI)。P < 0.05为差异有统计学意义。
2 结果 2.1 基因型分析每个DNA样本同时进行两管PCR分析,每管含一序列特异性引物(F1或F2),用以扩增有SNP特异性位点的目的片段。只有当序列特异性引物3' 端的碱基和模板DNA配对才可扩增出相应的特异性产物。若一样本同一位点F1和F2两条特异性引物均能扩增出产物,则表明其含有与两种引物相配对的碱基,为杂合子;若只能扩增出一种,则样本为纯合子。同时,每管含有的一对内参引物(C,R)均应扩增出产物,用以检验反应体系及条件的正确性。见图 1。

|
图 1 IL-17A基因SSP-PCR产物电泳图 上图为-692C/T位点条带:M:DNA marker,1:CT基因型,2:CC基因型,3:TT基因型,目的基因片段长度为290 bp,内参基因片段长度为427 bp;下图为-197G/A位点条带:M:DNA marker,1:GA基因型,2:GG基因型,3:AA基因型,目的基因片段长度为272 bp,内参基因片段长度为199 bp。 |
2.2 遗传平衡检验
哮喘组和健康对照组IL-17A基因两位点的基因型分布频率应用Hardy-Weinbergy遗传平衡定律检验,结果显示P > 0.05,达到遗传平衡,说明研究对象具有群体代表性。
2.3 PCR产物测序结果分析研究表明[4-5],应用SSP-PCR法检测SNPs位点与直接基因测序一样准确可靠,同时具有快速方便、经济实惠的优点。本研究随机选取的杂合子DNA样本SNPs的基因型测序结果与用SSP-PCR检测的结果是一致的,说明本研究应用SSP-PCR法检测基因位点的SNPs同样是准确可靠的。
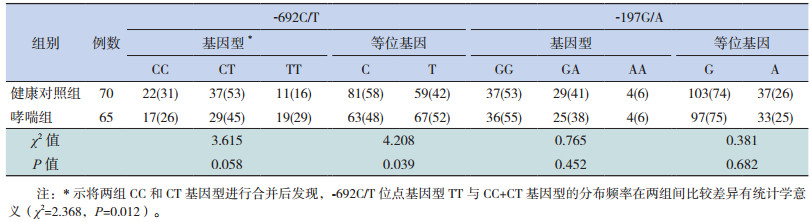
2.4 IL-17A-692C/T、-197G/A位点基因型及等位基因分布频率分析在哮喘组和健康对照组间,-692C/T位点基因型分布差异无统计学意义(P > 0.05),但将每组CC和CT基因型进行合并后比较发现,哮喘组TT基因型分布频率(29%)显著高于健康对照组(16%)(P=0.012)。哮喘组-692T等位基因分布频率(52%)显著高于健康对照组(42%)(P=0.039),且罹患儿童哮喘的风险T等位基因携带者是C等位基因携带者的1.413倍(OR=1.413,95%CI:1.015~1.917)。哮喘组和健康对照组间-197G/A位点基因型及等位基因分布频率差异均无统计学意义(P > 0.05)。见表 2。
| 表 2 哮喘组和对照组IL-17A基因型和等位基因分布频率[例(%)] |
3 讨论
哮喘是一种全球性疾病,在不同国家和地区均有分布,其患病率在1%~18%,严重地影响了人们的身心健康[6-8]。而哮喘的发病率和病死率也是逐年增加,据统计,目前以每10年20%~50%的速度增长[9]。同样,儿童哮喘也是严重影响小儿身心健康的最常见的呼吸道疾病。第三次(2009年9月至2010年8月)中国城市儿童哮喘流行病学调查显示(与2000年调查比较),我国儿童哮喘的现患率增加了50.6%,累积患病率增加了52.8% [10]。因此,对于哮喘致病机制的研究从而进行早期预防迫在眉睫,有极大的社会学和经济学意义。
虽然哮喘的发病率逐年增加,医学家及科学家们也从遗传与社会环境进行了深入的研究,但其发病机制仍不清楚。流行病学调查发现,儿童哮喘的发病率要远远高于成人。儿童喘息分为4种类型,而持续性喘息是最终能发展为哮喘的一类,该类患儿气道有典型的病理特征,其症状常常迁延不愈至成人阶段[11]。因此,早期哮喘风险度的评估对于这类儿童至关重要,若能进行有效的早期评估,进而对高风险儿童采取早期预防措施,相信能大大降低这类儿童哮喘的发病率。但遗憾的是,目前尚无合适的相关预测指标。据相关研究发现,哮喘的遗传率达到36~79%[12]。因此,对相关基因的检测可能是今后进行哮喘早期干预的重要手段。近年来,哮喘的分子遗传流行病学成为国际研究的一个热点,而其重点就是寻找哮喘的候选基因。目前报道的与哮喘发病相关的候选基因已超过200个[13-14]。
人IL-17家族有IL-17A至IL-17F 6个成员,IL-17主要由Th17细胞产生,而在中性粒细胞和嗜酸性粒细胞也有表达。IL-17与IL-17受体结合后可以通过JAKS/STATs信号途径促进各种因子(IL6、IL-8、粒细胞趋化蛋白2等)的转录[15]。而这些因子能聚集并激活各类炎症细胞,从而介导炎症瀑布反应,影响人体各种感染性、免疫性疾病的发生发展。有研究也表明在哮喘患者的血清中IL-17A的表达是明显升高的[1]。同时,IL-17A基因的-197G/A和-692C/T位点与多种免疫性疾病相关。目前,IL-17A启动子区-197G/A基因多态性与疾病易感性的相关性研究是分子遗传流行病学研究的热点,已有其与多种肿瘤(卵巢癌、胃癌等)、免疫相关性疾病(川崎病、炎症性肠病等)易感性相关的报道[16-17]。而在国内外的不同种族和地区都进行了针对-197G/A位点SNP与儿童哮喘易感性的研究,有阳性结果亦有阴性结果[18-19]。同时,检索文献发现目前尚无IL-17A基因-692C/T位点SNP与儿童哮喘易感性相关性研究的报道,但已有其与溃疡性结肠炎易感性相关的报道,本研究首次采用SSP-PCR法研究其基因多态性与儿童哮喘的关系。
本研究应用SSP-PCR检测了泸州地区儿童IL-17A -197位点和-692位点的SNP,通过统计分析发现:在本研究对象中IL17A基因启动子区-692C/T、-197G/A存在多态性;IL-17A基因启动子区-692位点的3个基因型在健康对照组儿童中分布频率情况:CC型31%,CT型53%,TT型16%,等位基因以C型(58%)为多;与Vargas-Alarcón等[20]报道的在墨西哥人群中CC型5.0%,CT型33.0%,TT型61.0%的分布情况有所不同,这可能与研究对象的地区和种族差异有关,这也能从侧面说明哮喘的发病与基因遗传相关,与地区和人种的差异有关;将哮喘组与对照组对比,IL-17A基因启动子区-692位点的TT基因型、T等位基因的分布频率差异有统计学意义(P < 0.05),提示IL-17A基因启动子区-692位点T等位基因可能是儿童哮喘发病的易感基因,携带T等位基因者罹患儿童哮喘的风险比携带C等位基因者高1.413倍。而在本研究中IL-17A基因-197G/A位点的基因型及等位基因分布频率在哮喘组和对照组间比较差异无统计学意义,提示IL-17A基因启动子区-197G/A位点基因多态性可能与儿童哮喘的易感性不相关。本研究结果将为儿童哮喘易感基因的生物信息数据库提供支持,进而为寻找到哮喘的候选基因,为患病高风险人群早期干预奠定基础。
综上,IL-17A基因启动子区-692位点SNP与儿童哮喘易感性相关,-692TT基因型携带者更易罹患哮喘。至于-692位点的SNP与IL-17A蛋白的表达是否相关,则需功能学试验的进一步研究。同时,基因多态性在不同区域同一族群,相同区域不同族群间的分布频率是不尽相同的,故有可能与疾病的易感性的关系也存在差异。所以,将研究对象扩展至不同区域、不同族群进行多中心大样本的采集研究,IL-17A基因多态性与儿童哮喘易感性的关系才能得到进一步的揭示。
| [1] | 吴红梅. 血清IL-17在支气管哮喘患儿中的表达意义[J]. 中国医药指南 , 2014, 12 (23) : 49–50. |
| [2] | Li J, Tian H, Jiang HJ, et al. Interleukin-17 SNPs and serum levels increase ulcerative colitis risk:a meta-analysis[J]. World J Gastroenterol , 2014, 20 (42) : 15899–15909. DOI:10.3748/wjg.v20.i42.15899 |
| [3] | 陈育智.支气管哮喘[M]//胡亚美, 江载芳.诸福棠实用儿科学.第7版.北京:人民卫生出版社, 2002:635. |
| [4] | 宋妙丽, 杨晓春, 申咏梅, 等. PCR-SSP法和基因测序检测人胰腺癌细胞株Patu 8988 K-ras基因第12位密码子点突变及其方式[J]. 苏州大学学报(医学版) , 2008, 28 (4) : 545–548. |
| [5] | 朱春燕, 聂咏梅, 周豪杰. PCR-SSP法检测HPA-15基因频率的可靠性分析[J]. 热带医学杂志 , 2015, 15 (7) : 897–898. |
| [6] | Losappio L, Heffier E, Contento F, et al. Thunderstorm-related asthma epidemic owing to Olea Europaea pollen sensitization[J]. Allergy , 2011, 66 (11) : 1510–1511. DOI:10.1111/all.2011.66.issue-11 |
| [7] | Kuschner WG. The asthma epidemic[J]. N Engl J Med , 2007, 356 (10) : 1073. DOI:10.1056/NEJMc063596 |
| [8] | Shafazand S, Colice G. Asthma:the epidemic has ended, or has it?[J]. Chest , 2004, 125 (6) : 1969–1970. DOI:10.1378/chest.125.6.1969 |
| [9] | Chen ZH, Wang PL, Shen HH. Asthma research in China:a five-year review[J]. Respirology , 2013, 18 (Suppl 3) : 10–19. |
| [10] | 全国儿科哮喘协作组, 中国疾病预防控制中心环境与健康相关产品安全所. 第三次中国城市儿童哮喘流行病学调查[J]. 中华儿科杂志 , 2013, 51 (10) : 729–735. |
| [11] | Akdis CA, Akdis M, Bieber T, et al. Diagnosis and treatment of atopic dermatitis in children and adults:European Academy of Allergology and Clinical Immunology/American Academy of Allergy, Asthma and Immunology/PRACTALL Consensus Report[J]. J Allergy Clin Immunol , 2006, 118 (1) : 152–169. DOI:10.1016/j.jaci.2006.03.045 |
| [12] | Vogt H, Bråbäck L, Zetterström O, et al. Asthma heredity, cord blood IgE and asthma-related symptoms and medication in adulthood:a long-term follow-up in a Swedish birth cohort[J]. PLoS One , 2013, 8 (6) : e66777. DOI:10.1371/journal.pone.0066777 |
| [13] | Zhou H, Hong X, Jiang S, et al. Analyses of associations between three positionally cloned asthma candidate genes and asthma or asthma-related phenotypes in a Chinese population[J]. BMC Med Genet , 2009, 10 : 123. DOI:10.1186/1471-2350-10-123 |
| [14] | Li X, Wills-Karp M, Ewart S. Investigating Gata3 as a positional candidate gene for allergic asthma in a murine model[J]. Int J Immunogenet , 2006, 33 (5) : 333–337. DOI:10.1111/eji.2006.33.issue-5 |
| [15] | Meyer DM, Jesson MI, Li X, et al. Anti-inflammatory activity and neutrophil reductions mediated by the JAK1/JAK3 inhibitor, CP-690, 550, in rat adjuvant-induced arthritis[J]. J Inflamm (Lond) , 2010, 7 : 41. DOI:10.1186/1476-9255-7-41 |
| [16] | Wang H, Zhang Y, Liu Z, et al. The IL-17A G-197A and IL-17F 7488T/C polymorphisms are associated with increased risk of cancer in Asians:a meta-analysis[J]. Drug Des Devel Ther , 2015, 9 : 5159–5168. |
| [17] | Long ZW, Yu HM, Wang YN, et al. Association of IL-17 polymorphisms with gastric cancer risk in Asian populations[J]. World J Gastroenterol , 2015, 21 (18) : 5707–5718. DOI:10.3748/wjg.v21.i18.5707 |
| [18] | Maalmi H, Beraies A, Charad R, et al. IL-17A and IL-17F genes variants and susceptibility to childhood asthma in Tunisia[J]. J Asthma , 2014, 51 (4) : 348–354. DOI:10.3109/02770903.2013.876647 |
| [19] | 王娟, 周娟, 蔺丽慧, 等. IL-17基因启动子区单核苷酸多态性与儿童哮喘的关系[J]. 第二军医大学学报 , 2011, 32 (5) : 481–484. |
| [20] | Vargas-Alarcón G, Angeles-Martínez J, Villarreal-Molina T, et al. Interleukin-17A gene haplotypes are associated with risk of premature coronary artery disease in Mexican patients from the Genetics of Atherosclerotic Disease (GEA) study[J]. PLoS One , 2015, 10 (1) : e0114943. DOI:10.1371/journal.pone.0114943 |