 2016, Vol. 18
2016, Vol. 18



癫癎是由多种病因引起的脑功能障碍综合征,是危害儿童身心健康和认知功能的神经系统疾病,目前癫癎的发病机制还不十分清楚 ,但多数学者认为遗传因素是一个重要的发病原因 [1]。在癫癎人群中约有 25%的患儿进展为难治性癫癎 [2]。谷氨酸受体基因(GRIK2)定位于 6号染色体 q21区域,研究表明该区域与多种精神疾病的易感性相关 [3]。GRIK2是离子型谷氨酸受体家族的一员,在大脑中大量表达,尤其是在海马和杏仁核区 [3]。Guo等 [4]研究人员发现 GRIK2基因与癫癎的易感性相关。由于环境、地域以及人群的差异,该基因的多态性是否与我国中部地区汉族人群中儿童癫癎的易感性相关值得进一步探讨。本研究以我国中部地区儿童癫癎患儿为研究对象,探讨 GRIK2基因两个 SNP位点(rs9390754和 rs4840200)的多态性与儿童癫癎的相关性。
1 资料与方法 1.1 研究对象选取 2009年 6月至 2014年 9月在我院就诊、诊断为癫癎的患儿 284例作为研究对象。患儿均为来自我国中部地区的汉族儿童。癫癎的诊断标准参考 2005年国际抗癫癎联盟(ILAE)对癫癎的定义 [5]:癫癎是一种脑部疾病,特点是持续存在产生癎性发作的持久性改变,并出现相应的神经生物学、认知、心理学及社会等方面的后果;诊断癫癎至少需要一次以上的癎性发作。
284例患儿中,难治性癫癎 132例,非难治性癫癎组 152例。其中难治性癫癎的诊断标准为:患儿每月发作 4次以上,正规使用抗癫癎药物且血药浓度在有效范围内,治疗两年以上仍不能控制发作,同时排除中枢性神经系统疾病或占位性病变 [6]。选取同期来我院体检的中部地区正常汉族儿童 315例作为对照。284例癫癎患儿中,男 160例,女 124例,平均年龄 5±3岁;对照组 315例儿童中,男 173例,女 142例,平均年龄 5±3岁。两组儿童的年龄、性别差异无统计学意义(P>0.05)。本研究得到我院医学伦理委员批准及患儿家长的知情同意。
1.2 基因组 DNA提取分别抽取癫癎患儿和对照组儿童静脉血2 mL,利用提基因组的试剂盒(上海生工公司)提取 DNA,并测定浓度。PCR分别扩增含有两个 SNP位点的片段。设计的引物分别为 F1:CAGTCTATGTTTTGACACTGAT,R1:CAATTAGAGCAGAGAACAGAAAAG(引入酶切位点 EcoRV,扩增含 rs9390754的片段);F2:CACAATTACATATGCACCATTGAATT,R2:AGAGGTTCA AGAACCCAAGAAGC(引入酶切位点 EcoRI,扩增含 rs4840200的片段)。PCR反应体系为 25 μL,含有 10倍浓度 PCR反应缓冲液 2.5 μL,10 mM dNTP混合液 1 μL,10 μM引物各 1 μL,Taq酶 1U(TAKARA公司 )。PCR反应条件:94℃ 3 min;94℃ 30 s,57℃ 30 s,72℃ 30 s,共 34个循环。PCR产物分别利用限制性内切酶 EcoRV和 EcoRI进行酶切分析。
1.3 统计学分析所有数据采用 SPSS 16.0统计软件分析,频率计数法计算基因型和等位基因频率,组间基因型频率分布和等位基因频率分布比较采用 χ2检验,P<0.05表示差异有统计学意义。
2 结果 2.1 癫癎组及对照组中 GRIK2基因型频率分布癫癎组和对照组两组的基因型均符合 HardyWeinberg遗传平衡定律(P>0.05),说明各组具有群体代表性。该基因 SNP位点 rs9390754的基因型确定:PCR产物经酶切电泳后 GG基因型显示 172 bp 1个条带,AA基因型显示 149 bp 1个条带,而 AG基因型显示以上两个条带。SNP位点rs4840200的基因型确定:经酶切以后 TT基因型显示 166 bp 1个条带,CC基因型显示 142 bp 1个条带,CT基因型显示以上两个条带。见图 1。

|
图 1 GRIK2 基因 SNP 位点的基因型电泳分析 A:rs9390754 位点的 3 种基因型:AA、AG 和 GG;B:rs4840200位点的 3 种基因型:CT、TT 和 CC。 |
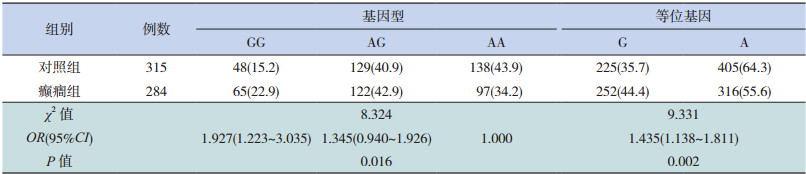
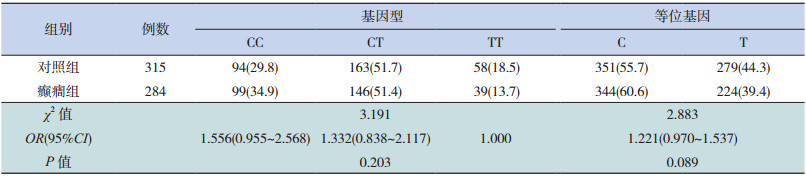
SNP位点(rs9390754)的基因型分布及等位基因分布与对照组相比差异均有统计学意义(P<0.05),见表 1。利用 PS(Power and SampleSize Program)软件对其群体的统计学效率分析得出 power值为 0.870,排除了假阳性的可能。而另一个 SNP位点(rs4840200)的基因型分布及等位基因分布与对照组相比差异均无统计学意义,见表 2。
| 表 1 GRIK2 基因 SNP 位点(rs9390754)基因型及等位基因在癫癎组及对照组中的分布[n(%)] |
| 表 2 GRIK2 基因 SNP 位点(rs4840200)基因型及等位基因在癫癎组及对照组中的分布[n(%)] |
2.2 GRIK2基因两个 SNP位点的基因型频率在癫癎亚组中的分布
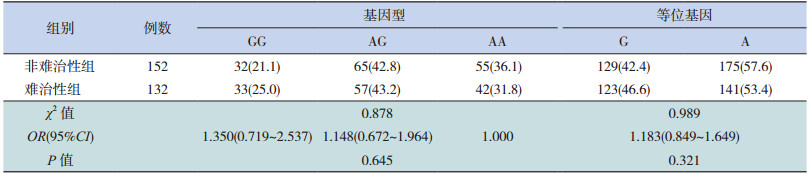
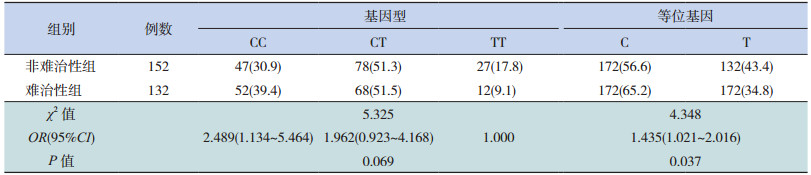
GRIK2基因的 SNP位点(rs9390754)的基因型频率以及等位基因频率在难治性癫癎和非难治性癫癎两组间差异无统计学意义(分别 P=0.645、0.321),见表 3。SNP位点(rs4840200)的基因型频率在两组之间差异无统计学意义,但难治性癫癎组患儿携带 C等位基因的频率显著高于非难治性癫癎组患儿(P=0.037),见表 4。
| 表 3 GRIK2 基因 SNP 位点(rs9390754)基因型频率在癫癎亚组中的分布[n(%)] |
| 表 4 GRIK2 基因 SNP 位点(rs4840200)基因型频率在癫癎亚组中的分布[n(%)] |
3 讨论
谷氨酸是中枢神经系统内重要的兴奋性神经递质,在突触的信号传递、学习记忆以及神经系统发育过程中发挥着重要作用;而谷氨酸受体分为两类:代谢型和离子型受体 [7]。GRIK2是离子型受体家族的一员,GRIK2基因位于 6q21区,也就是存在于被报道的多种精神疾病的易感连锁区域。2015年 Manuel等 [8]利用全基因组关联分析(GWAS)发现该基因与强迫性的神经失调存在相关性。Cordoba等 [9]研究发现 GRIK2基因的异常突变可能导致智力障碍等相关的疾病。Lee等 [10]研究小组发现 GRIK2基因的三联碱基 TAA的重复突变与亨廷顿舞蹈症的发病年龄不存在相关性。
Griswold等 [11]和 Casey等 [12]均发现 GRIK2基因可能与孤独症谱系障碍的发病相关。GRIK2一直被认为是癫癎的候选基因,细胞水平改变 GRIK2基因的表达实验以及敲除的小鼠模型实验均表明该基因对癫癎发作的维持以及控制存在相关性 [13]。SNP位点(rs9390754)位于GRIK2基因的内含子区域,本研究发现该位点的多态性与癫癎的易感性相关,G等位基因为风险因子,携带 GG基因型的人群患癫癎的风险是 AA基因型的 1.927倍,该结论与 Guo等 [4]研究小组的结果具有一致性,推测该位点可能单独或者连锁其他位点共同影响 GRIK2基因的表达水平,导致机体在兴奋传导过程中发生异常,进而导致癫癎的发作,具体的分子机制还需进一步研究证实。
SNP位点 rs4840200也位于 GRIK2基因的内含子区域,本研究显示该位点的多态性与癫癎的易感性不相关,该结论与 Guo等 [4]研究小组的结果存在差异,可能的原因是:(1)地区及人群不同;(2)本研究的研究对象均是儿童患者,不包括成人,而 Guo等 [4]的研究对象包括成人。
此外,本研究根据治疗效果将癫癎患儿分为难治性癫癎和非难治性癫癎两个亚组,并比较各位点基因型在两组间的分布差异,发现 SNP位点(rs4840200)的 C等位基因频率在两组中的分布差异有统计学意义,推测该位点可能与癫癎的疗效相关,即携带 C等位基因的患儿发作不容易控制,易进展为难治性癫癎。
综上所述,GRIK2基因的 SNP位点(rs9390754)的多态性与癫癎的易感性相关,而另一 SNP位点(rs4840200)的 C等位基因可能是容易发生耐药的遗传学危险因素,该结论对深入认识癫癎的发病及耐药机制提供了一定的依据。但以上结论需要大样本、多中心的进一步验证,具体的分子机制还需要今后进一步研究。
| [1] | Hani AJ, Mikait HM, Mikait MA. Genetics of pediatric epilepsy[J]. Pediatr Clin North Am,2015, 62 (3) :703–722 . |
| [2] | 孟曙庆, 张洪. 难治性癫癎的治疗进展[J]. 中华临床医师杂志,2013,7 (15) :7115–7119. |
| [3] | 高成阁, 陈元堂, 余冰, 等. 谷氨酸受体基因多态性与精神分裂症的相关性研究[J]. 中华行为医学与脑科学杂志,2009,18 (12) :1093–1096. |
| [4] | Guo YL, Larry W, Sham PC, et al. Two-stage genome-wide association study identifies variants in CAMSAP1L1 as susceptibility loci for epilepsy in Chinese[J]. Hum Mol Genet,2012, 21 (5) :1184–1189 . |
| [5] | Fisher RS, van Emde Boas W, Blume W, et al. Epileptic seizures and epilepsy: definitions proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE)[J]. Epilepsia,2005, 46 (4) :470–472 . |
| [6] | 吴逊, 沈鼎烈. 难治性癫癎[J]. 中华神经科杂志,1998,31 (1) :4. |
| [7] | Laqranha VL, Matte U, Carvalho TG, et al. Increased glutamate receptor and transporter expression in the cerebral cortex and striatum of gcdh-/- mice: possible implications for the neuropathology of glutaric acidemia type I[J]. PLoS One,2014, 9 (3) :e90477. |
| [8] | Manuel M, Samuels JF, Wang Y, et al. Genome-wide association study in obsessive-compulsive disorder: results from the OCGAS[J]. Mol Psychiatry,2015, 20 (3) :337–344 . |
| [9] | Cordoba M, Rodiguez S, Gonzalez MD, et al. Expanding the spectrum of Grik2 mutations: intellectual disability, behavioural disorder, epilepsy and dystonia[J]. Clin Genet,2015, 87 (3) :293–295 . |
| [10] | Lee JH, Lee JM, Ramos EM, et al. TAA repeat variation in the GRIK2 gene does not influence age at onset in Huntington's disease[J]. Biochem Biophys Res Commun,2012, 424 (3) :404–408 . |
| [11] | Griswold AJ, Ma D, Cukier HN, et al. Evaluation of copy number variations reveals novel candidate genes in autism spectrum disorder-associated pathways[J]. Hum Mol Genet,2012, 21 (15) :3513–3523 . |
| [12] | Casey JP, Maqalhaes T, Conroy JM, et al. A novel approach of homozygous haplotype sharing identifies candidate genes in autism spectrum disorder[J]. Hum Genet,2012, 131 (4) :565–579 . |
| [13] | Telfeian AE, Federoff HJ, Leone P, et al. Overexpression of GluR6 in rat hippocampus produces seizures and spontaneous nonsynaptic bursting in vitro[J]. Neurobiol Dis,2000, 7 (4) :362–374 . |